The 'place cells', something like our brain GPS
Orientation and exploration in new or unknown spaces is one of the cognitive faculties that we use most often. We use it to guide us in our house, our neighborhood, to go to work.
We also depend on it when we travel to a new and unknown city for us. We use it even when we drive and, possibly, the reader will have been the victim of a carelessness in his orientation or in that of a companion, which will have condemned him to be lost, being forced to turn around with the car until he with the appropriate route.
It's not the orientation's fault, it's the hippocampus's fault
All these are situations that often frustrate us and that lead us to curse our orientation or that of others with insults, shouts and various behaviors. All right, because today I will give a brushstroke in the neurophysiological mechanisms of orientation , in our Brain GPS to understand us.
We will start by being specific: we should not curse the orientation since it is only a product of our neural activity in specific regions. Therefore, we will begin by cursing our hippocampus.
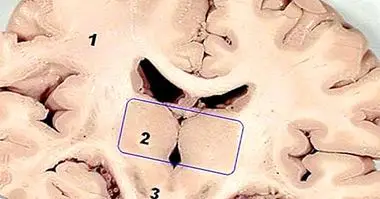
The hippocampus as a brain structure
Evolutively, the hippocampus is an ancient structure, it is part of the arquiculture, that is, those structures that are phylogenetically older in our species. Anatomically, it is part of the limbic system, in which other structures such as the amygdala are also found. The Limbic System is considered the morphological substrate of memory, emotions, learning and motivation.
The reader possibly if he is accustomed with psychology will know that the hippocampus is a necessary structure for the consolidation of declarative memories, that is, with those memories with episodic content about our experiences or else, semantic (Nadel and O'Keefe, 1972) .
Proof of this are the abundant studies that exist about the popular case of the "patient HM", a patient whose temporal hemispheres had been removed, producing a devastating anterograde amnesia, that is, he could not memorize new facts although he retained most of the of your memories from before the operation. For those who want to deepen in this case I recommend the studies of Scoville and Millner (1957) who studied the HM patient exhaustively.
The Place Cells: what are they?
So far we do not say anything new, or anything surprising. But it was in 1971 when by chance a fact that generated the beginning of the study of navigation systems in the brain was discovered. O'keefe and John Dostrovski, using intracranial electrodes, could record the activity of hippocampal-specific neurons in rats . This offered the possibility that while performing different behavioral tests, the animal was awake, conscious and moving freely.
What they did not expect to discover was that there were neurons that responded selectively depending on the area in which the rat was. It is not that there were specific neurons to each position (there is no neuron for your bathroom, for example), but that they were observed in the CA1 (a specific region of the hippocampus) cells that marked reference points that could be adapted to different spaces .
These cells were called place cells. Therefore, it is not that there is a neuron of place for each specific space that you frequent, but rather they are points of reference that relate you to your environment; This is how egocentric navigation systems are formed. Place neurons will also form allocentric navigation systems that will relate elements of the space between them.
Innate programming vs. experience
This discovery perplexed many neuroscientists who considered the hippocampus as a declarative learning structure and now saw how it was able to encode spatial information. This gave rise to the hypothesis of the "cognitive map" that would postulate that a representation of our environment would be generated in the hippocampus.
Just as the brain is an excellent generator of maps for other sensory modalities such as the coding of visual, auditory and somatosensory signals; it is not unreasonable to think of the hippocampus as a structure that generates maps of our environment and that guarantees our orientation in them .
The research has gone further and has put this paradigm to the test in very different situations. It has been seen, for example, that the cells of place in the maze tasks shoot when the animal makes mistakes or when it is in a position in which the neuron would usually shoot (O'keefe and Speakman, 1987).In tasks in which the animal must move through different spaces it has been seen that place neurons shoot depending on where the animal comes from and where it is going (Frank et al., 2000).
How spatial maps are formed
Another of the main focuses of research interest in this field has been on how these spatial maps are formed. On the one hand we could think that place cells establish their function based on the experience we receive when we explore an environment, or, we might think that it is an underlying component of our brain circuits, that is, innate. The question is not yet clear and we can find empirical evidence that supports both hypotheses.
On the one hand, the experiments of Monaco and Abbott (2014), which recorded the activity of a large number of cells, have seen that when an animal is placed in a new environment several minutes pass until these cells start to shoot with normal. So that, place maps would be expressed, in some way, from the moment an animal enters a new environment , but the experience would modify these maps in the future.
Therefore, we might think that brain plasticity is playing a role in the formation of spatial maps. Then, if plasticity really played a role, we would expect that knockout mice to the NMDA receptor of the neurotransmitter glutamate - that is, mice which do not express this receptor - would not generate spatial maps because this receptor plays a fundamental role in brain plasticity and learning.
Plasticity plays an important role in the maintenance of spatial maps
However, this is not the case, and it has been seen that knockout mice to the NMDA receptor or mice that have been treated pharmacologically to block this receptor, express similar patterns of response of the cells in new or familiar environments. This suggests that the expression of spatial maps is independent of brain plasticity (Kentrol et al., 1998). These results would support the hypothesis that navigation systems are independent of learning.
In spite of everything, using logic, the mechanisms of cerebral plasticity must be clearly necessary for the stability in the memory of recently formed maps. And, if it were not so, what would be the use of the experience that one forms by walking the streets of his city? Would not we always have the feeling that it is the first time we entered our house? I believe that, as on so many other occasions, the hypotheses are more complementary than they seem and, in some way, despite an innate functioning of these functions, plasticity has a role to play in maintaining these spatial maps in memory .
Network, address and edge cells
It is quite abstract to talk about place cells and possibly more than one reader has been surprised that the same brain area that generates memories serves us, so to speak, GPS. But we are not finished and the best is yet to come. Now let's curl the curl really. Initially, it was thought that space navigation would depend exclusively on the hippocampus when it was seen that adjacent structures such as the entorhinal cortex showed very weak activation as a function of space (Frank et al., 2000).
However, in these studies the activity in ventral areas of the entorhinal cortex was recorded and in later studies, dorsal areas were recorded, which have a greater number of connections to the hippocampus (Fyhn et al., 2004). So that it was observed that many cells of this region fired depending on the position, similar to the hippocampus . So far they are expected to find results but when they decided to increase the area they would register in the entorhinal cortex they had a surprise: among the groups of neurons that were activated depending on the space occupied by the animal there were apparently silent zones - that is, they were not activated-. When regions that showed activation were virtually joined, patterns were observed in the form of hexagons or triangles. They called these neurons of the entorhinal cortex "red cells".
When red cells were discovered, it was possible to solve the question of how cells are formed. Having the cells place numerous connections of the network cells, it is not unreasonable to think that they are formed from them. However, once again, things are not so simple and experimental evidence has not confirmed this hypothesis. The geometric patterns that form the network cells have not been able to be interpreted yet.
Navigation systems are not reduced to the hippocampus
The complexity does not end here. Even less when it has been seen that navigation systems are not reduced to the hippocampus. This has made it possible to expand the limits of research to other brain areas, thus discovering other cell types related to the cells of place: Steering cells and edge cells .
Steering cells would code the direction in which the subject moves and would be located in the dorsal tegmental nucleus of the brainstem. On the other hand, edge cells are cells that increase their firing rate as the subject approaches the limits of a given space and can be found in the subiculum-specific region of the hippocampus. We are going to offer a simplified example in which we will try to summarize the function of each type of cell:
Imagine that you are in the dining room of your house and that you want to go to the kitchen. Since you are in the dining room of your house, you will have a room cell that will fire while you stay in the dining room, but since you want to go to the kitchen you will also have another activated room cell that represents the kitchen. The activation will be clear because your house is a space that you know perfectly and the activation can be detected both in the cells of place and in the cells network.
Now, start walking towards the kitchen. There will be a group of specific address cells that will now be firing and will not change as long as you maintain a specific direction. Now, imagine that to go to the kitchen you must turn right and cross a narrow corridor. The moment you turn, your address cells will know it and another set of address cells will register the direction that it has now taken to activate, and the previous ones will be deactivated.
Imagine also that the corridor is narrow and any false movement can cause you to hit the wall, so your edge cells will increase your firing rate. The closer you get to the corridor wall, the higher the firing ratio would show your edge cells. Think of the edge cells as the sensors that some new cars have and that make an audible signal when you are maneuvering to park. The edge cells They work in a similar way to these sensors, the closer they are to colliding the more noise they make . When you arrive at the kitchen, your place cells will have told you that it has arrived satisfactorily and as it is a wider environment, your edge cells will relax.
Let's just complicate everything
It is curious to think that our brain has ways to know our position. But there is still a question: How do we reconcile declarative memory with space navigation in the hippocampus ?, that is, how do our memories influence these maps? Or could it be that our memories were formed from these maps? To try to answer this question we must think a little further. Other studies have pointed out that the same cells that code space, of which we have already spoken, also code the time . Thus, there has been talk of time cells (Eichenbaum, 2014) which would code the perception of time.
The surprising thing about the case is that more and more evidence supporting the idea that place cells are the same as time cells . Then, the same neuron using the same electrical impulses is able to code space and time. The relationship between the coding of time and space in the same action potentials and their importance in memory remain a mystery.
In conclusion: my personal opinion
My opinion about it? Taking off my scientist's robe, I can say that the human being is accustomed to thinking about the easy option and we like to think that the brain speaks the same language that we . The problem is that the brain offers us a simplified version of the reality that he himself processes. In a way similar to the shadows of Plato's cave. So, just as in quantum physics barriers of what we understand as reality are broken, in neuroscience we discover that in the brain things are different from the world that we consciously perceive and we must have a very open mind that things do not have why be as we really perceive them.
The only thing that I have clear is something that Antonio Damasio is used to repeating a lot in his books: the brain is a great map generator . Maybe the brain interprets time and space in the same way to map our memories. And if it seems chimerical to you think that Einsten in his theory of relativity one of the theories he postulated was that time could not be understood without space, and vice versa. Undoubtedly unraveling these mysteries is a challenge, even more so when they are difficult aspects to study in animals.
However, no effort should be spared on these issues. First of curiosity. If we study the expansion of the universe or the recently recorded gravitational waves, why would not we study how our brain interprets time and space? And, secondly, many of the neurodegenerative pathologies such as Alzheimer's disease have space-time disorientation as first symptoms.Knowing the neurophysiological mechanisms of this coding, we could discover new aspects that will help to better understand the pathological course of these diseases and, who knows, discover new pharmacological or non-pharmacological targets.
Bibliographic references:
- Eichenbaum H. 2014. Time cells in the hippocampus: a new dimension for mapping memories. Nature 15: 732-742
- Frank LM, Brown EN, Wilson M. 2000. Trajectory encoding in the hippocampus and entorhinal cortex. Neuron 27: 169-178.
- Fyhn M, Molden S, Witter MP, Moser EI, Moser M-B. 2004. Spatial representation in the entorhinal cortex. Science 305: 1258-1264
- Kentros C, Hargreaves E, Hawkins RD, Kandel ER, Shapiro M, Muller RV. 1998. Abolition of long-term stability of new hippocampal place cell maps by NMDA receptor blockade. Science 280: 2121-2126.
- Monaco JD, Abbott LF. 2011. Modular realignment of grid cell activity as a basis for hippocampal remapping. J Neurosci 31: 9414-9425.
- O'Keefe J, Speakman A. 1987. Single unit activity in the mouse hippocampus during a spatial memory task. Exp Brain Res 68: 1-27.
- Scoville WB, Milner B (1957). Loss of recent memory after bilateral hippocampallesion. J Neurol Neurosurg Psychiatry 20: 11-21.